New SPRm 220 Cell-based SPR
Newest commercial SPR Microscopy Label-free binding affinity of membrane proteins Higher throughput


BI-4500 & BI-2500 SPR series
Prestigious 2016 R&D 100 Award winner 5 & 3 microfluidic channels Benchtop SPR

Leader in Electrochemistry SPR
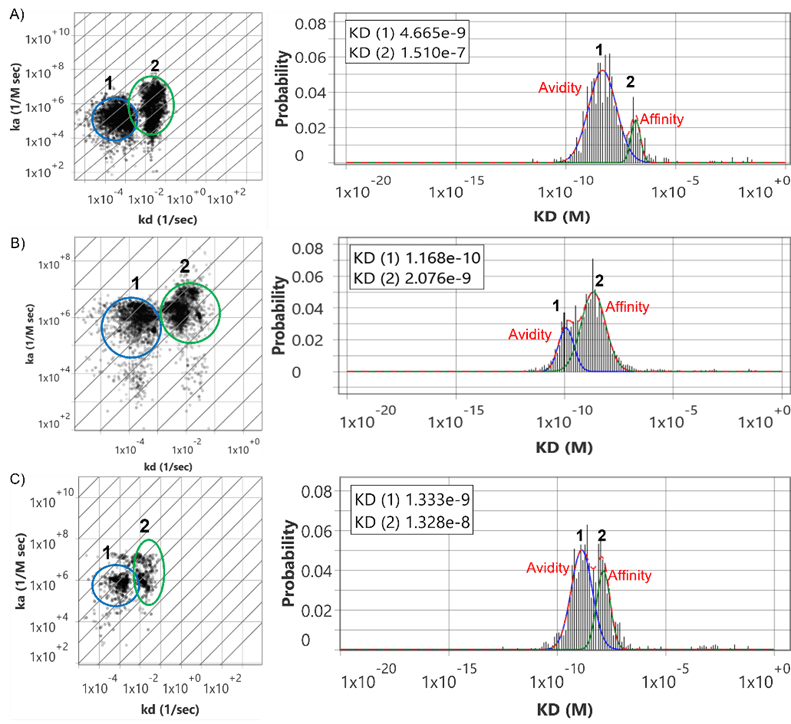
Over 20 years publishing with EC-SPR Conformational changes of proteins Studies in reorganization of organic thin films
Biosensing Instrument Offers Bioanalytical Tools & Measurement Service To Accelerate Drug Discovery Research
Leading innovation


Enabling discovery


SPR MICROSCOPY
CELL-BASED BINDING ASSAY
SPRm 220 system seamlessly integrates optical microscopy with SPR to enable a label-free, real-time and in-vitro measurement of binding reactions and kinetics at membranes of individual cells.
INTRODUCTION OF SPRM

This video introduces the SPR Microscopy technology, and how it is used to detect small molecules and their interaction with membrane protein in vitro.
SPR TECHNOLOGY
MODULAR AND COST-EFFECTIVE
Our modular designs are tailored to your individual research needs, all at an affordable price. BI’s unique analysis modules allow for a research focus on life science, electrochemical enhanced SPR, and gas SPR.
HOW SPR WORKS

This popular video explains what SPR technology is, how it is used to detect small molecules and their interaction with other proteins.
NEWS
Posted on Sep 17 2025
Biosensing Instrument will be at Discovery on Target 2025
Posted on Aug 19 2025
Measurement Services Promotion Information
Posted on May 19 2025
Biosensing Instrument will be at Drug Discovery Chemistry 2025
Posted on Mar 04 2025
Biosensing will be at BPS 2025!
Posted on Feb 03 2025
Versatile Applications
LIFE SCIENCE & CELL-BASED ASSAY

ELECTROCHEMICAL MEASUREMENTS

CHEMICAL VAPOR SENSING

ENVIRONMENT, FOOD & BEYOND