Most membrane proteins carry some form of glycans, which regulate many cellular processes such as cell differentiation and cell-to-cell interactions.1 There are two types of glycans based on the type of linkage to specific amino acids on proteins: N-linked and O-linked glycans.2 Cancer cells, including pancreatic cancer, have aberrant expression of both types of glycans on membrane proteins.3&4 Therefore, understanding the changes that occur in the glycosylated cancerous environment is critical to rational drug design.
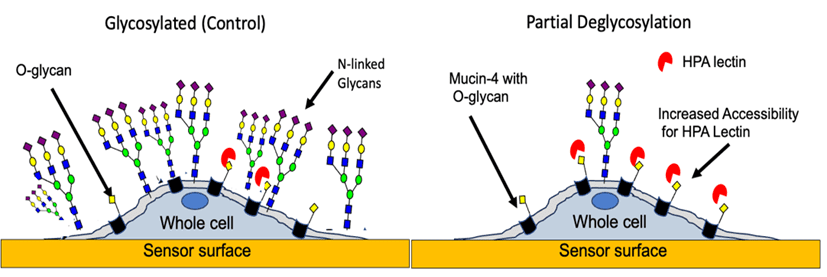
Figure 1: Pancreatic cancer cells on the sensor surface model for O-glycan profiling with HPA under glycosylated and after the partial removal of N-glycans.
Lectins, such as Helix Pomatia Agglutinin (HPA), are predominantly used to identify and profile specific sugar epitopes of O-glycans.5 Also, HPA can recognize multiple O-glycan epitopes and has been widely used in cancer biomarker discovery.6 In general, variations in binding interactions resulting from simultaneous binding of lectin to multiple receptors carrying glycans, different antigen glycoforms, and receptor accessibility are all collectively referred here as “heterogeneity”.
The capability to determine binding heterogeneity as well as real-time binding kinetics simultaneously is a unique feature of surface plasmon resonance microscopy (SPRM). This approach allows for a more accurate understanding of the “true” binding interactions of new or potential therapeutic drugs.7
In this study, O-linked glycan binding interactions and their underlying heterogeneity under N-linked glycosylated and partial N-linked deglycosylated conditions in native pancreatic cancer (BXPC3)cells were studied using HPA (Figure 1).8 Pancreatic cancer (PC) cells were used in this study as they exhibit aberrant N-linked and O-linked glycosylation compared to normal pancreatic cells.8 Also, enzymatic N-linked deglycosylation of whole pancreatic cancer cells using PNGase enzyme allowed testing of the hypothesis that aberrant N-linked glycosylation in and around membrane proteins play a major role in hindering their binding interactions.
The kinetic results obtained using SPRm 200 instrument identified three predominant binding interaction modes for HPA lectin in glycosylated BxPC3 cells, which produced KDs mentioned in Table 1. Interestingly, as seen in Figure 2A, a faint fourth binding mode (mode d) was also identified, which was not as abundant as the other three modes with a low probability of 0.014. These different binding modes of interactions for HPA could be identified by varying both the sample concentration and measurement regions within a single cell or among different cells.
| BXPC3 cells | KD (nM) | 95% CI (nM) | ka (M-1s-1) | kd (s-1) |
| Glycosylated | a) 1.2 b) 59 c) 0.82 d) 0.022 | a) 1.1 to 1.3 b) 51 to 60 c) 0.77 to 0.84 d) 0.019 to 0.023 | a) 6.1X106 b) 2.0X105 c) 4.3X 105 d)7.0X106 | a) 7.4X10-3 b) 1.0X10-2 c) 3.5X10-4 d) 1.5X10-4 |
| Deglycosylated | a) 0.58 b) 10 c) 0.25 e) 4.5 | a) 0.51 to 0.61 b) 9 to 11 c) 0.22 to 0.26 e) 4.3 to 5 | a) 6.2X105 b) 9.2X105 c) 2.0 X107 e) 2.4X104 | a) 3.6X10-4 b) 1.0X10-2 c) 2.5X10-3 e) 1.1X10-4 |
Table 1: Kinetic parameters of HPA lectin binding to BxPC3 cells. All data are representative from 5 different experiments. A 1:2 binding model was applied to all the data analysis.
The influence of N-linked glycosylation in BXPC3 cells was studied by partially deglycosylating the cells with the enzyme PNGase F. A total of four binding interaction modes upon removal of N-linked glycans became more apparent relative to the glycosylated control, and their binding kinetics were determined. Under deglycosylated conditions, a fourth mode (e) of binding interaction is clearly presented, having a significantly lower on-rate and weaker affinity than modes a and c (Table 1) and, possibly indicating that this mode is particularly sensitive to steric hindrance as seen in Figure 2B. Modes a, b, and c shifted to slightly higher affinities, respectively. Additionally, this study identified a faint mode (mode d) with high affinity that interestingly disappears after deglycosylation, possibly inferring that N-linked glycans might also play a limited role in enhancing some modes of HPA interactions as seen in Figure 2B.
Kinetic parameters were extracted from hundreds of sensorgrams directly measured from the binding interactions of single cells from of numerous sensor chips. These results suggest that N-linked glycosylation significantly influences the HPA interaction by not only hindering access but also by completely inhibiting one of the binding modes (mode e), which becomes more readily accessible after N-linked deglycosylation. Clearly, HPA appears to have multiple presentations of its O-glycan epitopes for binding interactions.9&10 Although the identity of the specific O-glycoforms HPA recognizes in this study are unknown, the binding kinetics of the most prominent HPA epitopes under glycosylated and partial deglycosylated conditions have been determined using SPRM.
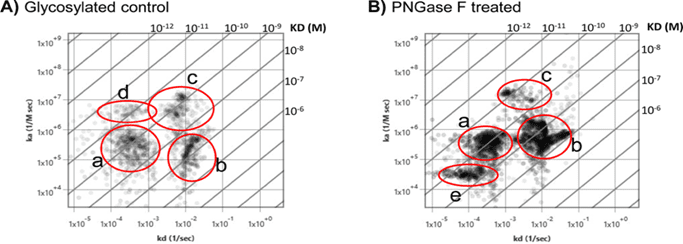
Figure 2: Removal of N-linked glycans unmasks HPA binding heterogeneity. A) Affinity isotherm plot extracted from hundreds of responsive ROIs showing four distinct HPA binding population modes on glycosylated BxPC3 cells. B) Affinity isotherm plot extracted from hundreds of responsive ROIs for HPA on deglycosylated BxPC3 cells showing four distinct binding populations. Mode e highlights the appearance of a new binding population mode upon enzymatic deglycosylation. a, b, c, d, and e represent binding interaction modes.
This study has identified multiple interaction modes for O-glycoprotein binding kinetics on PC cells, demonstrating that in vitro whole-cell binding kinetic studies without disrupting the native environment of membrane glycoproteins are critical for revealing more biologically relevant binding interaction information. This level of sub-cellular resolution on binding heterogeneity would not be readily observable using traditional end-point analysis or other low-resolution approaches which typically average the binding responses over multiple cells in an experiment.
Author: Jesús Aguilar Díaz de león, Nguyen Ly, and Miyuki Thirumurthy | Biosensing Instrument | Published February 5th, 2025
DOWNLOAD PDF
Download a PDF of Application Note 162: N-linked Glycan Deglycosylation Unmasks New HPA O-linked Glycan Heterogeneity
- Costa, Ana Filipa, et al. "Targeting glycosylation: a new road for cancer drug discovery." Trends in cancer 6.9 (2020): 757-766.
- Pinho, Salomé S., and Celso A. Reis. "Glycosylation in cancer: mechanisms and clinical implications." Nature Reviews Cancer 15.9 (2015): 540-555.
- Fuster MM, Esko JD (2005) The sweet and sour of cancer: Glycans as novel therapeutic targets. Nat Rev Cancer 5:526–542. pmid:16069816.
- Stowell S. R., Ju T., & Cummings RD (2015) Protein Glycosylation in Cancer. Annu Rev Pathol 10:473–510 pmid:25621663.
- Guzman-Aranguez A, Argüeso P (2010) Structure and biological roles of mucin-type O-glycans at the ocular surface. Ocular Surface 8:. pmid:20105403.
- Dwek M V., Ross HA, Streets AJ, Brooks SA, Adam E, Titcomb A, et al. (2001) Helix pomatia agglutinin lectin-binding oligosaccharides of aggressive breast cancer. Int J Cancer 95:. pmid:11241316.
- Han C, Dong T, Wang P, Zhou F (2022) Microfluidically Partitioned Dual Channels for Accurate Background Subtraction in Cellular Binding Studies by Surface Plasmon Resonance Microscopy. Anal Chem 94:. pmid:36454605.
- Aguilar Díaz de león, Jesús S., Miyuki Thirumurthy, and Nguyen Ly. "Surface plasmon resonance microscopy identifies glycan heterogeneity in pancreatic cancer cells that influences mucin-4 binding interactions." Plos one 19.5 (2024): e0304154.
- Springer G. F. (1989) Tn epitope (N-acetyl-D-galactosamine alpha-O-serine/threonine) density in primary breast carcinoma: a functional predictor of aggressiveness. Mol Immunol 26:1–5 pmid:2467192.
- Rambaruth ND, Greenwell P, Dwek M V. (2012) The lectin Helix pomatia agglutinin recognizes O-GlcNAc containing glycoproteins in human breast cancer. Glycobiology 22:. pmid:22322011